
Alelopatía

Introducción
Liberación de aleloquímicos
Relaciones plantas/ambiente
La ecología vegetal estudia las relaciones entre las plantas y su ambiente, incluidas otras plantas. Esas relaciones consisten en factores abióticos y bióticos que afectan a las plantas, y los efectos de las plantas sobre esos factores.La disponibilidad de luz, la temperatura, las condiciones de humedad, las diferentes características del suelo, como la disponibilidad de nutrientes y otros organismos, son los factores más importantes (Crawley, 1997).
Modifica el ambiente
Los efectos de una planta incluyen alteraciones en esos factores en la vecindad de la planta. Una planta puede cambiar la disponibilidad de luz debajo de su follaje, puede afectar la calidad y cantidad de nutrientes disponibles para la planta, y puede alterar muchos otros procesos en su vecindad.
A otros organismos
En particular, una planta puede combatir (o atraer) otras plantas, patógenos, herbívoros y depredadores que pueden dañarla o beneficiarla a ella o a la vegetación vecina.
Planta viva
Los procesos pueden ocurrir a medida que la planta vive o después de su muerte a medida que avanza la descomposición. Los compuestos químicos se pueden producir y liberar durante cualquiera de estos procesos, y pueden mostrar actividad alelopática en las interacciones planta-planta (Rice, 1984).
Planta muerta
Coevolucionado con microorganismos
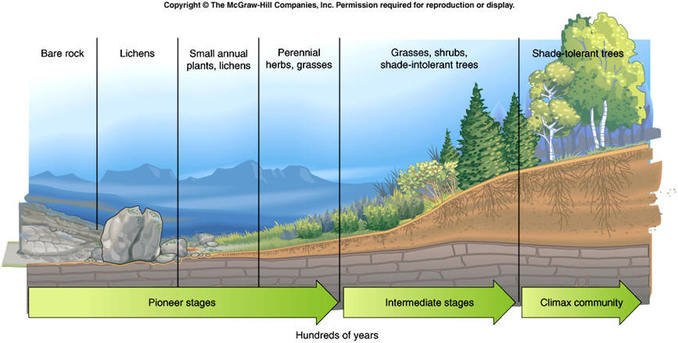
Colonizan áreas recién expuestas
Las comunidades de invertebrados y microbios generalmente colonizan las superficies de tierra recién expuestas antes de los autótrofos, y por lo tanto se convierten en el primer conjunto comunitario establecido en áreas desnudas (Hodkinson et al. 2002).
Establecimiento de plantas
Esas comunidades heterotróficas conservan nutrientes que facilitan el establecimiento de plantas autótrofas. Por lo tanto, es probable que durante cientos de millones de años, las plantas terrestres superiores hayan vivido constantemente en estrecho contacto con los microorganismos del suelo.
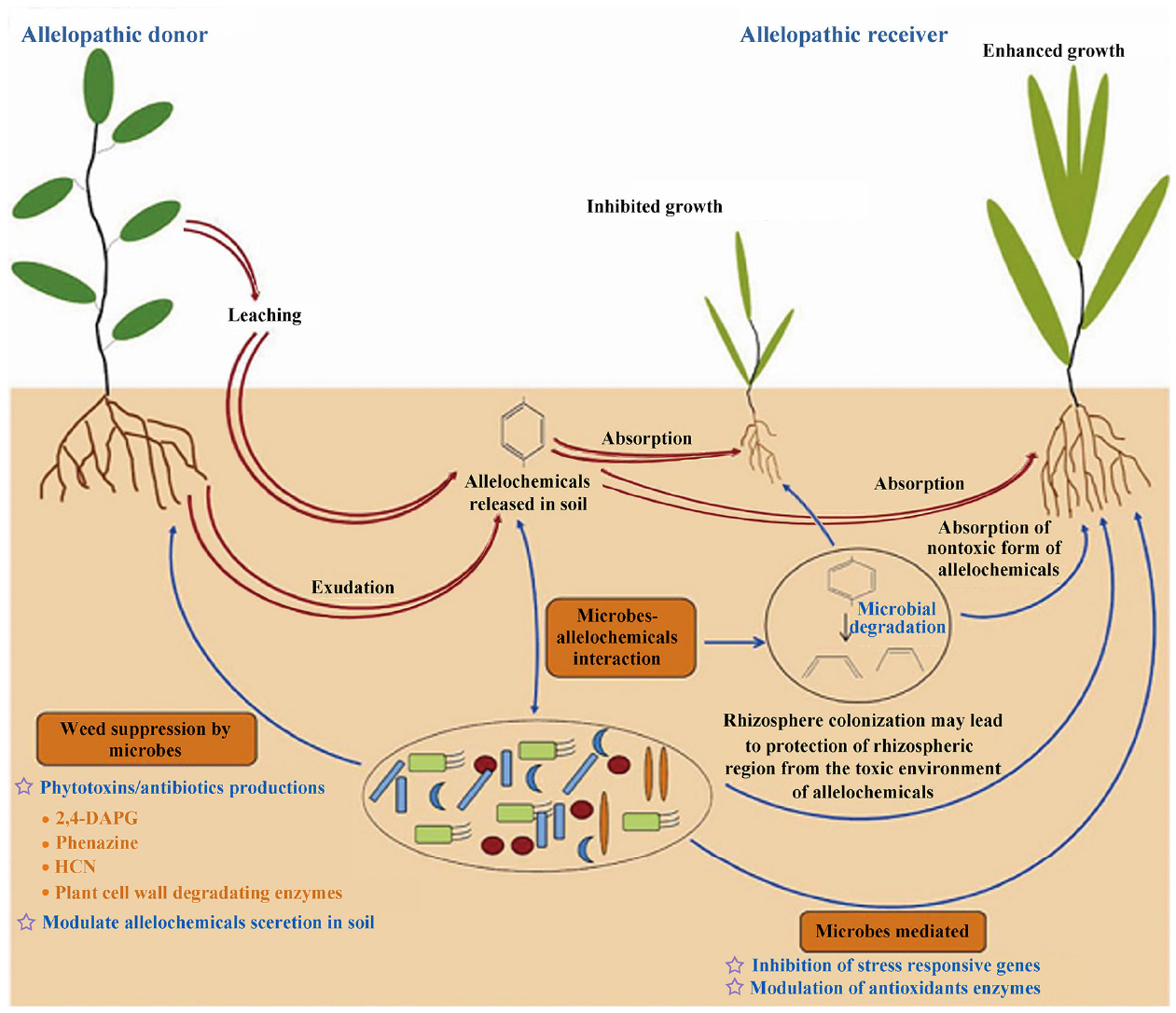
Liberación de fitoquímicos
Por lo tanto, cada proceso que libera fitoquímicos del tejido vegetal al medio del suelo ha evolucionado, mientras que los fitoquímicos liberados han sido influidos por la actividad microbiana.La degradación microbiana puede haber neutralizado, reducido o amplificado cualquier efecto fitotóxico de una planta en otra planta (Williamson y Weidenhamer, 1990).

Microorganismos/Plantas
Sin embargo, las bacterias del suelo no son competidores superiores ni para los aleloquímicos ni para los nutrientes: las plantas pueden pero las bacterias no pueden almacenar nutrientes para su uso posterior; y las poblaciones bacterianas del suelo están reguladas de manera efectiva por la depredación.Por estas razones, cantidades ecológicamente significativas de aleloquímicos pueden haber sido capaces de afectar recurrentemente las células de la raíz de las plantas receptoras incluso en comunidades de plantas con coevolución natural (Williamson y Weidenhamer, 1990; Wardle y Lavelle, 1997; Mallik, 2002).
Degradación bacteriana de aleloquímicos
Como resultado de esta coexistencia entre las plantas y los microorganismos del suelo, cualquier especie que libere aleloquímicos debe haber evolucionado en presencia de la degradación bacteriana de los aleloquímicos liberados en su comunidad vegetal original.
Probabilidad de degradación rápida de fitoquímicos
Además, la planta puede haber evolucionado para minimizar la probabilidad de que las especies receptoras y los microorganismos del suelo puedan desintoxicar rápidamente los compuestos liberados.
Tolerar y utilizar fitoquímicos
Los microbios abundantes en una comunidad de plantas, por otro lado, deberían haber evolucionado para tolerar y utilizar los fitoquímicos que se liberan de las especies de plantas con las que han evolucionado (ver Schmidt et al. 2000).
Otras plantas
Aunque se ha argumentado que la actividad microbiana en el suelo conduce inevitablemente a la inactivación de todos los agentes alelopáticos antes de que afecten a las especies receptoras, hoy en día existe evidencia que sugiere que varias especies de plantas no relacionadas han evolucionado para desintoxicar aleloquímicos originados de plantas en la interfaz suelo-raíz (ver más abajo).
Dos diferentes perspectivas
Evidencias
Tradicionalmente se ha considerado que la prueba inequívoca de la existencia de alelopatía requiere (ver Fuerst y Putnam, 1983; Blum et al. 1999):
Carrera armamentista
En un contexto ecológico y evolutivo, apenas es necesario suponer que todos estos pasos siempre permanecen distinguibles una vez evolucionados. Si la alelopatía ha sido evolutivamente significativa para ambas contrapartes de la interacción, y si el objetivo de la especie donante es inhibir el crecimiento de la especie receptora, debería haber una carrera armamentista evolutiva entre estos dos.
Condiciones y etapas fenológica
El resultado de la carrera puede depender drásticamente de las condiciones ambientales, y puede cambiar a medida que evolucionan las nuevas adaptaciones. Dependiendo de la etapa de la variedad, las especies objetivo pueden sufrir daños de una manera ecológicamente significativa, o pueden no verse perjudicadas en absoluto.
Dos proceso separados y dos objetivos
Por lo tanto, dos procesos separados y dos objetivos diferentes son distinguibles de los cuatro pasos presentados anteriormente. El problema en cuestión puede abordarse utilizando la perspectiva de estudiar una especie donante o la perspectiva de estudiar una especie receptora (Koricheva y Shevtsova, 2002; Reigosa et al. 2002).
Objetivos
Perspectiva de la especie receptora
Si reacciona
Cuando se usa la perspectiva de la especie receptora, es relevante probar si una especie receptora potencial reacciona a la existencia de aleloquímicos potenciales en su vecindad, en su superficie o en ella, y si la respuesta de la especie objetivo es ecológicamente significativa.
Relevancia de la respuesta
La especie receptora puede responder alterando el crecimiento o desintoxicando los compuestos mediante un proceso químico que no se utiliza para otros fines (Einhellig, 1996; Weidenhamer, 1996; Schulz y Wieland, 1999). La respuesta de crecimiento proporciona evidencia directa de que la alelopatía es actualmente ecológicamente significativa, mientras que la última respuesta sugiere fuertemente que la alelopatía ha afectado la evolución de las especies receptores. Además, la última respuesta puede requerir recursos que no pueden asignarse a otra parte, lo que puede afectar la aptitud de la planta (ver Crawley, 1997).
Especie donante en la actualidad
Si se observa cualquiera de estas respuestas, puede que no siempre sea necesario demostrar que en la actualidad existe una especie donante que es capaz de liberar agentes alelopáticos en cantidades efectivas dentro de la distribución actual de la especie receptora, debido a la tolerancia de la receptora. Las especies, las condiciones de crecimiento o la distribución de especies pueden haber cambiado después de que la respuesta evolucionó. Obviamente, esto no justifica especulaciones poco realistas sobre lo que pudo haber ocurrido en el pasado (véase Romeo, 2000).
Perspectiva de la especie donante
Liberación de fitoquímicos
Otro proceso totalmente separado es utilizar la perspectiva de la especie donante (Recuadro 1). Aquí, es relevante demostrar que una especie donante potencial libera fitoquímicos debido a la presencia de una especie receptora probable, o que la liberación de compuestos químicos es inducida por la presencia de una especie receptora potencial.
Especie receptora probable o potencial
Las plantas donantes pueden distinguir la presencia de plantas receptoras al reconocer fitoquímicos específicos producidos por estas (Sunderland, 1960; Logan y Stewart, 1992), o simplemente pueden reaccionar ante la fluctuación ambiental en condiciones abióticas (por ejemplo, un cambio en el pH del suelo) que es generalmente causado por una planta receptora; muchas especies de plantas tienden a cambiar las condiciones abióticas alrededor de sus raíces (Waisel et al. 1996).
Liberación continua o recurrente
En una estrategia alternativa, una especie donante potencial libera de forma continua o recurrente compuestos que tienen (o tuvieron) un efecto significativo en una especie receptora que interfiere con la especie donante en circunstancias naturales. En tal caso, la fluctuación abiótica en las condiciones ambientales puede regular la toxicidad de la liberación (Muller y del Moral, 1966; Nilsson et al. 1998).
Éxito de la especie donante
La evolución de las especies receptoras puede o no haber sido afectada por los compuestos liberados, dependiendo de cuán importante haya sido la presión selectiva causada por los compuestos alelopáticos durante la evolución de las especies receptoras. Obviamente, la alelopatía puede haber sido crucial para el éxito de una especie donante, y completamente irrelevante para el éxito de su especie receptora.

Planta receptora
Relaciones ecológicas y diseños experimentales
Es importante distinguir la diferencia entre las relaciones ecológicas y los diseños experimentales relacionados con las plantas receptoras de los relacionados con las plantas donantes. Las relaciones anteriores se han explorado en la literatura ecológica en los últimos años. Los ecólogos de plantas han utilizado con éxito cámaras radiculares, carbón activado y diferentes densidades de plantas receptoras para afirmar que varias especies deben hacer frente a la alelopatía en situaciones naturales y estimar las implicaciones de la alelopatía en las plantas receptoras (Mahall y Callaway, 1992; Nilsson, 1994 ; Thijs et al.1994; Sinkkonen, 2001).
centaurea VS gramíneas
Callaway y Aschehoug (2000) encontraron que en la vecindad de una centaurea euroasiática [centaurea difusa (Centaurea diffusa Lamarck)] el crecimiento de especies de gramíneas de América del Norte disminuye más que el crecimiento de especies de gramíneas estrechamente relacionadas de comunidades nativas de C. diffusa. Además, el crecimiento de las raíces de una de las especie receptora de América del Norte [Festuca idahoensis Elmer] fue más lento en contacto con las raíces de C. diffusa, pero el efecto disminuyó cuando el suelo se trató con carbón activado (Ridenour y Callaway , 2001).La razón más probable para el fenómeno observado son las sustancias inhibidoras, cuya concentración se diluye gradualmente a medida que aumenta la distancia desde las raíces en contacto (tenga en cuenta que el carbón activado per se no excluye la posibilidad de que los fitoquímicos exudados inmovilicen cantidades significativas de nutrientes en las proximidades de las raíces de los donantes, y que la proporción de nutrientes inmovilizados disminuye gradualmente a medida que aumenta la distancia de las raíces de los donantes en los tratamientos sin carbono).

Centaurea diffusa

Festuca idahoensis
Diseños experimentales dependientes de la densidad
Otra forma elegante de dilucidar la existencia de alelopatía es planear un diseño experimental dependiente de la densidad en el que las plantas receptoras crezcan a diferentes densidades en el suelo que contenga una concentración homogénea de posibles sustancias aleloquímicas encontradas naturalmente.
Capacidad de desintoxicación
Las relaciones ecológicas relacionadas con una especie receptora pueden abordarse con un estrategia alternativa: para probar si las especies receptoras iniciales son capaces de desintoxicar posibles aleloquímicos, y afirmar que la capacidad de desintoxicación no ha evolucionado debido a otros factores ecológicos.
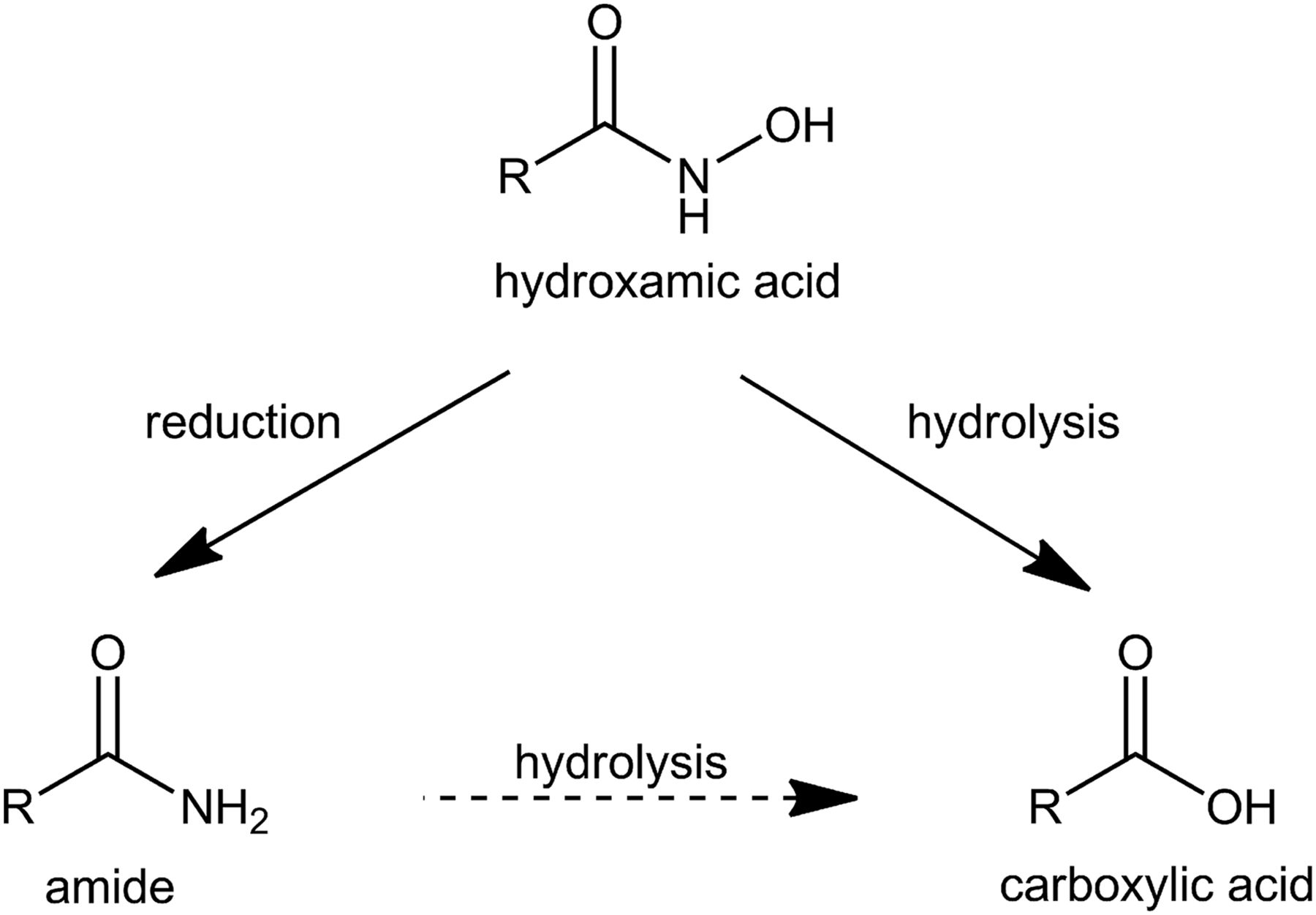
Ácidos hidroxámicos
Las desintoxicaciones de los productos de degradación de los ácidos hidroxámicos en muchas especies de plantas dicotiledóneas parecen demostrar la utilidad de esta estrategia. Los ácidos hidroxámicos son químicos de defensa generalizados en las Poaceae (Niemeyer, 1988). Son bien conocidos por sus múltiples funciones en la resistencia a plagas y enfermedades y por su actividad alelopática contra otras plantas (Niemeyer, 1988; Chiapusio et al. 1997; Frey et al. 1997). La benzoxazolinona es un metabolito fitotóxico y abundante de ácidos hidroxámicos. La capacidad de desintoxicación de la benzoxazolinona varía considerablemente entre las especies dicotiledóneas relacionadas, dependiendo de la frecuencia probable de contacto con los pastos productores de ácido hidroxámico en la historia evolutiva de la especie (Schulz y Wieland, 1999). Es probable que varias especies de plantas hayan evolucionado para desintoxicar agentes alelopáticos que se originan en miembros liberadores de ácido hidroxámico de Poaceae, o pueden haber evolucionado para utilizar microbios de rizosfera que desintoxican los agentes alelopáticos. Dado que la benzoxazolinona no se produce en el suelo independientemente de las plantas productoras de ácido hidroxámico (o sus residuos), este caso especial proporciona evidencia confiable de la importancia de las interacciones alelopáticas en circunstancias naturales.

Poaceas

Dicotiledóneas
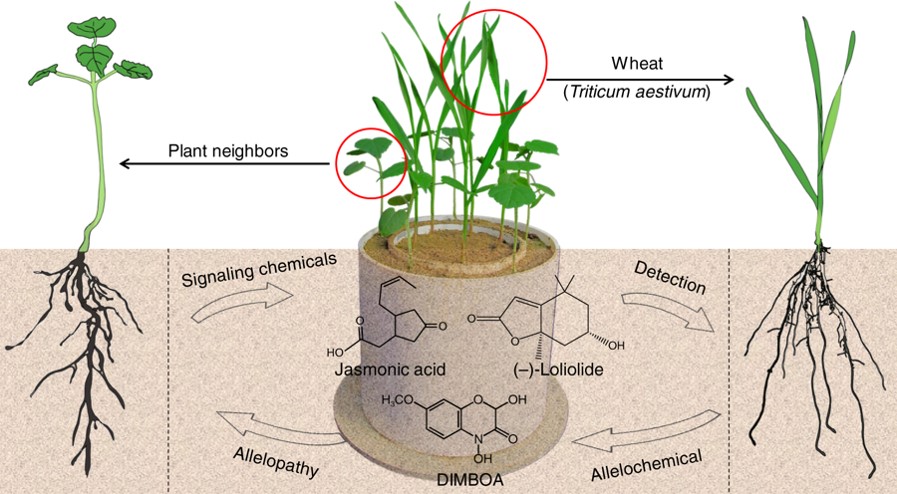
Planta donante
Por el contrario de las relaciones de las especies receptoras, rara vez se han estudiado las relaciones ecológicas de las especies donantes.
Planta libera fitoquímicos
Para demostrar que la alelopatía es evolutivamente significativa, uno debería ser capaz de demostrar que una planta libera, sintetiza o asigna fitoquímicos para afectar el crecimiento de otras plantas (ver Siemens et al. 2002), no solo que los compuestos liberados tienen efectos tóxicos sobre otras plantas en condiciones ambientales adecuadas (ver Seigler, 1996; Romeo, 2000). El último fenómeno siempre puede ser un efecto secundario ecológicamente insignificante (o incluso una compensación) de una función primaria que ha sido significativa en la evolución de las especies donantes
Crecimiento de otras plantas
Función alelopática
Si las causalidades no son claras, a menudo es fácil negar el papel evolutivo significativo de la función alelopática si los aleloquímicos potenciales tienen funciones concurrentes en la ecología de una especie donante (ver Pellissier, 1995; Brierley et al. 2001).
Estrategia evolutiva estable (ESS)
Debido a estas razones, podría ser valioso estudiar si la liberación de agentes alelopáticos es una estrategia evolutiva estable (ESS) para las especies donantes potenciales (ver Maynard-Smith, 1972, 1989). Una ESS es una estrategia que es adoptada por la mayoría de la población y que no puede ser desplazada por un mutante raro (un ejemplo hipotético podría ser una mayor exudación de aleloquímicos específicos a través de puntas de raíz a parches de nutrientes versus exudación inalterada de aleloquímicos específicos a través de puntas de raíz a nutrientes parches).

Relaciones ecológicas
Este enfoque evolutivo se ha utilizado con éxito para explicar las relaciones ecológicas en diferentes entornos. Por ejemplo, las interacciones alelopáticas entre diferentes especies de algas pueden explicar cómo pueden coexistir múltiples especies en un ambiente aparentemente uniforme (Czárán et al. 2002), y la alelopatía de dos especies de plantas terrestres que interfieren puede permitir la coexistencia de estas (Dubey y Hussain, 2000 ; Amarasekare, 2002).
En la perspectiva
La diferencia entre la perspectiva de estudiar una planta donante y la perspectiva de estudiar una planta receptora es tan grande como la diferencia entre las perspectivas de estudiar un herbívoro y estudiar una planta que consume el herbívoro (Koricheva y Shevtsova, 2002). En la literatura ecológica reciente, muchos estudios que abordan los metabolitos secundarios de las plantas y las interacciones planta-herbívoro examinan el problema en cuestión utilizando cualquiera de las perspectivas, no ambas (ver Hartley y Jones, 1997). Si se abordan simultáneamente, las dos perspectivas se separan conscientemente (ver Alliotta y Branca, 1996). Sin embargo, por alguna razón, un uso mixto de dos perspectivas diferentes era común cuando se abordaban los metabolitos secundarios de las plantas y las interacciones planta-planta en el siglo XX. Esto puede haber causado parcialmente el sesgo de la investigación de la alelopatía hacia preguntas no orientadas ecológicamente (ver Bever et al. 1998; Pellissier, 1998; Watkinson, 1998; Carral Vilariño, 2002).
Efectos alelopáticos
Tres posibilidades
Esquema traducido de Sinkkonen (2006)Las interacciones alelopáticas ecológicamente significativas de los metabolitos secundarios originados en plantas se pueden dividir en tres categorías en circunstancias naturales, en función de la historia evolutiva de las especies donantes (Figura 1). Primero, los productos químicos liberados para fines no alelopáticos pueden reducir o mejorar la aptitud de las plantas receptoras no intencionales, o una liberación involuntaria de fitoquímicos, por ejemplo debido a daños físicos, puede provocar alteraciones en el crecimiento de otras plantas. Esta categoría se puede llamar alelopatía no intencional. En segundo lugar, los productos químicos podrían liberarse para varios propósitos ecológicamente relevantes, uno de los cuales es la alelopatía contra otras plantas. Esta categoría se puede describir como alelopatía paralela. La alelopatía paralela puede ser el resultado de una alelopatía no intencional recurrente que ha llevado a una adaptación en la especie donante. La alelopatía paralela puede ser espacial o temporal: el mismo compuesto puede usarse para diferentes propósitos en diferentes partes de una planta, o en diferentes momentos, respectivamente. Tercero, los productos químicos podrían liberarse principalmente o únicamente debido a su actividad alelopática contra otras plantas. Esta alelopatía primaria puede no ser tan común como las otras categorías, ya que los fitoquímicos a menudo tienen múltiples funciones en la ecología de las plantas (ver Seigler y Price, 1976; Hartley y Jones, 1997; Nitao et al. 2002). Las relaciones ecológicas relacionadas con estas tres categorías difieren entre sí, y pueden abordarse utilizando diferentes diseños experimentales. En todos los casos, las plantas objetivo o donante, o ambas, se han adaptado o posiblemente se adaptarán a la presión selectiva causada por los aleloquímicos liberados.
Alelopatía no intencional
Por coincidencia
Las condiciones ambientales pueden cambiar para que los fitoquímicos se liberen en grandes cantidades, persistan más tiempo en el suelo o se degraden menos que antes (An et al. 2002). Una mutación o una serie de mutaciones pueden cambiar la calidad [por ejemplo, la adición de un grupo metoxi al esqueleto de bibencilo en crowberry (Empetrum nigrum L.); ver Nilsson et al. 2000] o cantidad [por ejemplo, diferencias en la cantidad de isotiocianatos en la canola (Brassica napus L.); ver Choesin y Boerner, 1991] de químicos liberados.Como resultado, los fitoquímicos pueden convertirse en aleloquímicos efectivos contra otras plantas.
Nuevo entorno
Una planta puede crecer en un entorno nuevo donde los efectos de los fitoquímicos liberados no son los mismos que en su entorno original. Los cambios en las relaciones alelopáticas de una planta donante pueden ocurrir si la planta ha colonizado una nueva comunidad de plantas (Schulz y Wieland, 1999) o una nueva área geográfica (Callaway y Aschehoug, 2000). Allí, los microbios del suelo pueden degradar los fitoquímicos liberados de las especies exóticas de manera menos eficiente que dentro de su distribución nativa, o la vegetación indígena puede carecer de la capacidad de tolerar los nuevos fitoquímicos.
Sistemas agrícolas
La alelopatía no intencional puede ocurrir en sistemas agrícolas cuando el hombre reúne especies de cultivos y malezas con historias evolutivas separadas (Schulz y Wieland, 1999).
Paralela o primaria
Si la alelopatía no intencional aumenta la aptitud de la planta donante, el fenómeno puede volverse recurrente. Finalmente, el resultado será una alelopatía paralela o primaria.Por el contrario, si una especie receptora no intencional se ve afectada esporádicamente por fitoquímicos liberados de otra especie por cualquier razón, la probabilidad de adaptación seguirá siendo baja (ver Gardner et al. 1998).
Germinación en cajas Petri
La prueba de germinación en caja Petri de lechuga es un ejemplo clásico de un diseño experimental que evalúa el potencial de un compuesto para engendrar alelopatía no intencional. En este experimento, las semillas de lechuga (Lactuca sativa L.) pueden germinar en papel de filtro a diferentes concentraciones de fitoquímicos de una especie no agrícola. La germinación de las semillas se registra regularmente y finalmente se mide el tamaño de las plántulas. El resultado generalmente indica que los compuestos seleccionados muestran potencial alelopático a ciertas concentraciones contra la germinación de lechuga y el crecimiento de plántulas en placas Petri (ver Inderjit y Dakshini, 1995; Inderjit y Weston, 2000).

Lactuca sativa
Hipótesis coevolutiva de Rabotnov

Comunidades nativas
La hipótesis coevolutiva de Rabotnov establece que es probable que la alelopatía sea menos significativa en comunidades de plantas coevolucionadas que en el caso de una especie nativa y exótica (Rabotnov, 1974; Mallik, 2002; Reigosa et al. 2002).
Plantas receptoras
Se puede considerar que la hipótesis de Rabotnov establece que la posibilidad de efectos alelopáticos drásticos es mayor en la alelopatía no intencional que en el caso de especies receptoras intencionadas, o que las especies receptoras eventualmente ganarán la carrera armamentista entre donantes y especies receptoras, posiblemente con la ayuda del suelo microbios.
Contradicciones
Además, si los aleloquímicos de
Especies donantes
Actualmente hay evidencia que contradice la hipótesis de Rabotnov en ciertas comunidades de plantas, o al menos sugiere que las especies donantes pueden tener éxito en la carrera durante un período de tiempo evolutivamente significativo (Mallik, 2002). A pesar de esto, puede pasar un tiempo antes de que el patrón de liberación de aleloquímicos de una especie donante exótica alcance un ESS en una nueva área geográfica, y antes de que las especies de plantas y microorganismos nativos se adapten a la nueva presión selectiva.
Roles no alelopáticos
las especies exóticas tenían múltiples roles defensivos dentro del área de distribución original de la especie, la importancia de otros que no sean roles alelopáticos puede disminuir en la nueva área si las especies exóticas escapan de sus plagas nativas cuando ingresa a la nueva área.
Alelopatía paralela
Importancia
Los capítulos anteriores de este libro han descrito la gran diversidad de fitoquímicos que están potencialmente involucrados en la alelopatía. Los compuestos descritos a menudo tienen múltiples funciones, y la importancia de las diferentes funciones de muchos fitoquímicos aún no se ha explorado.
Fitoquímicos
Causas variación
Compuestos fenólicos
Por ejemplo, no está claro cuáles son las principales causas de variación en los niveles de compuestos fenólicos en las plantas. Tradicionalmente, se ha pensado que los niveles fenólicos están mediados por las interacciones planta-herbívoro, pero los fenólicos también protegen a las plantas del fotodaño, afectan la disponibilidad de nutrientes en ciertas comunidades de plantas y son uno de los principales grupos de posibles agentes alelopáticos (Northup et al. 1995; Blum et al.1999; Close y McArthur, 2002; Nitao et al.2002).
Comportamiento de plantas donantes
Cuando se trata de alelopatía paralela, debe tenerse en cuenta que el comportamiento de las plantas donantes es siempre un arreglo entre la función alelopática y las funciones no alelopáticas de los fitoquímicos. Por lo tanto, las respuestas óptimas de las plantas donantes a la presencia de plantas receptoras pueden no encontrarse siempre, y las plantas receptoras pueden ser capaces de hacer frente a los compuestos liberados en cierta medida (Schulz y Wieland, 1999)
Función alelopatica / no alelopática
Fitoquímicos
Alelopatía primaria
La evidencia actual sugiere fuertemente que muchos metabolitos secundarios de las plantas son como las manos de una ardilla: han evolucionado para múltiples propósitos. A pesar de esto, la alelopatía primaria se puede encontrar en la naturaleza.
Producción o liberación

Plantas parásitas
La alelopatía primaria es a menudo ecológicamente significativa en las interacciones entre plantas parásitas y sus huéspedes (Rice, 1984).
Interacciones ecológicas
Efectos diluidos
Típicamente, la respuesta de un organismo a una toxina es la estimulación a bajas concentraciones (Figura 2). A medida que aumenta la concentración, la estimulación se convierte gradualmente en inhibición. A concentraciones muy altas, el organismo puede morir. El efecto de los aleloquímicos en las plantas generalmente sigue este patrón (Carballeira et al. 1988; An et al. 1993). En ciertos casos, la estimulación no se ha observado a bajas concentraciones, pero en teoría podría ocurrir si la concentración utilizada fuera lo suficientemente baja (An et al. 1993; Sinkkonen, 2001)
Estimulación a bajas concentraciones
Dosis de la interferencia química
Debido a la dependencia de la dosis de la interferencia química, las plantas donantes inevitablemente enfrentan problemas cuando intentan optimizar la liberación de aleloquímicos en un entorno fluctuante.
Ambientes fluctuantes
Densidad de la planta receptora
Además, debido a que las semillas de varias especies generalmente germinan cerca unas de otras en la naturaleza, las especies receptoras sensibles pueden beneficiarse de sus vecinos insensibles y abundantes que inactivan la mayoría de los aleloquímicos a altas densidades de plantas (ver Thijs et al. 1994)
Concentraciones más altas
Dado que el equilibrio entre la estimulación alelopática y la inhibición depende de la densidad de la planta receptora y la concentración fitoquímica en el suelo, las especies receptoras pueden tender a evolucionar hacia la estimulación a concentraciones fitoquímicas más altas (Sinkkonen, 2001).
Bajas concetraciones por altas densidades
Sin embargo, un posible costo de esta tendencia son los efectos estimulantes débiles a bajas concentraciones y a altas densidades de plantas. Por lo tanto, la tolerancia de una especie receptora puede incluso disminuir, si las densidades de las plantas son permanentemente altas o las concentraciones fitoquímicas son bajas (Figura 2).
Tolerancia
Plantas vecinas insensibles
Además, debido a que las semillas de varias especies generalmente germinan cerca unas de otras en la naturaleza, las especies receptoras sensibles pueden beneficiarse de sus vecinos insensibles y abundantes que inactivan la mayoría de los aleloquímicos a altas densidades de plantas (ver Thijs et al. 1994)
Material en descomposición
Densidad
Los efectos alelopáticos del material vegetal en descomposición también dependen de la densidad.
Aleloquímicos potenciales
Los residuos de plantas frescas por lo general contienen potenciales productos químicos alelopáticos, y las especies donantes pueden incluso haber evolucionado para utilizar las propiedades tóxicas de sus propios residuos para asegurar la supervivencia de la descendencia (Wardle et al. 1998).
Liberación de aleloquímicos
Lixiviación o inactivación
A medida que los microorganismos comienzan el proceso de descomposición, los aleloquímicos a menudo se liberan del material vegetal muerto y luego se lixivian por la lluvia o se metabolizan a compuestos inactivos, dependiendo de las condiciones abióticas (An et al. 1996; An et al. 2002).
Periodos estimulación / inhibición
El proceso a menudo provoca la inhibición del crecimiento de las plantas receptoras intencionadas o no, pero los residuos de algunas especies de plantas tienden a estimular el crecimiento de las plantas resultantes (Wardle et al. 1991b; An et al. 1996). A menudo hay un corto período de estimulación al comienzo del proceso de descomposición, seguido de un período inhibitorio más largo que gradualmente se convierte nuevamente en estimulación al final del proceso de descomposición de residuos (An et al. 1996).
Comparten aleloquímicos
Sin embargo, dependiendo de la densidad, las plantas receptoras pueden compartir los aleloquímicos para que no ocurra la inhibición (Figura 4).
Selección de receptores
Estrategias diversas
Dado que es probable que todas las especies receptoras no desarrollen estrategias similares para poder crecer bajo la influencia de agentes alelopáticos liberados de una sola especie donante,
Estrategia óptima
la especie donante debe encontrar una estrategia óptima para afectar negativamente el crecimiento de las plantas que interfieren tan eficientemente como sea posible en un entorno fluctuante continuo e inesperado.Si un ESS se logra solo liberando aleloquímicos, debería ser la media de las estrategias óptimas contra cada especie receptora abundante, gran plasticidad fenotípica en la liberación de aleloquímicos de las especies donantes.
Especies receptoras nativas
Debido a que diferentes especies de plantas utilizan diferentes microhábitats (Crawley, 1997; Miller y Bowman, 2003), y dado que ciertas especies de plantas cambian más probablemente las condiciones de crecimiento desfavorables para las especies donantes potenciales que otras especies de plantas (Ohlson et al. 2001), las especies receptoras nativas pueden haber sido de diferente importancia durante la evolución de la alelopatía en una especie donante.
Condiciones desfavorables para la donante
Efectos más severos
Como resultado, se puede esperar que los efectos alelopáticos de las especies donantes sean más severos contra aquellas especies receptoras que han tenido un alto potencial para disminuir la aptitud de las especies donantes en situaciones naturales. Esto puede explicar por qué una sola especie donante puede tener efectos fatales contra ciertas especies y efectos alelopáticos débiles o nulos contra otras especies receptoras potenciales (ver Wardle et al. 1998; Quested et al. 2003).
Choques repentinos VS estrés constante
Vecinas impredecibles
Las plantas son vecinas desagradables: son impredecibles. El tamaño, la edad y el vigor de las plantas donantes y receptoras determinan de manera sustancial la cantidad de productos químicos aleloquímicos que se pueden liberar y la cantidad que se puede tolerar.
Múltiples tensiones simultáneas
Múltiples tensiones simultáneas pueden modificar la respuesta de las plantas (Hartley y Jones, 1997), y la interferencia química puede afectar sinérgicamente con otros factores que causan estrés.
Espacial y temporal
Existe una variación espacial y temporal sustancial en la concentración de aleloquímicos y en el patrón de liberación de aleloquímicos entre especies e individuos, e incluso dentro de los individuos (Tabla 1).
Ambiente irregular
Debido a esto, las plantas donantes y receptoras a menudo viven en un entorno irregular. Este parche natural no descalifica la importancia de varios mecanismos de dilución o selección de receptores; simplemente complica la evaluación de la importancia de las interacciones alelopáticas.
Liberación de aleloquímicos
Razones
La variación relacionada con la liberación de aleloquímicos puede seleccionarse por varias razones.Primero, las comunidades de microorganismos del suelo parecen poder aclimatarse y adaptarse a concentraciones crónicas de aleloquímicos, lo que obviamente aumenta la degradación (Blum, 1996; Schmidt et al. 2000; Staman et al. 2001).En segundo lugar, la evidencia de los estudios de pesticidas de plantas sugiere que las especies receptoras desarrollan rápidamente una alta resistencia si los compuestos tóxicos están presentes de forma continua o repetida a altas concentraciones en el medio ambiente (Gardner et al. 1998). La evolución de la resistencia puede retrasarse o no ocurrir contra las fitotoxinas artificiales si las toxinas se liberan con poca frecuencia y en cantidades variables (Gardner et al. 1998). Esta estrategia también puede minimizar los costos de producir una toxina.Tercero, varias toxinas diferentes que afectan simultáneamente pueden retrasar la prevalencia de genotipos resistentes (Gardner, 2002). Por lo tanto, si las mezclas de aleloquímicos que se alternan cualitativa y cuantitativamente están presentes de manera irregular, la aclimatación y la adaptación a los aleloquímicos pueden retrasarse.
Choques repentinos
Por lo tanto, las condiciones bióticas pueden favorecer la estrategia de choques repentinos, en lugar de la estrategia de estrés constante en la liberación de aleloquímicos.
Conflicto de intereses
An y colaboradores (2001) estudiaron la actividad biológica de todos los aleloquímicos identificados como residuos de vulpia [Vulpia myuros (L.) C. C. Gmelin].
Aleloquímicos de vulpia

Vulpia myuros
Mayoría: compuestos actividad baja o media
Descubrieron que la mayoría de los compuestos poseían una actividad biológica baja o media y estos compuestos también contribuían con la mayor parte de la fitotoxicidad de la vulpia, mientras que los compuestos con alta actividad biológica eran minoritarios y solo estaban presentes en baja concentración (An et al. 2001).
Cóctel óptimo
También descubrieron que si se usaban aleloquímicos en proporciones encontradas en el residuo, la inhibición del crecimiento en las especies objetivo era mayor que si se usaran los mismos aleloquímicos en proporciones iguales. Por lo tanto, al menos algunas especies donantes pueden liberar "un cóctel óptimo" de aleloquímicos, lo que proporciona la mejor relación de entrada-salida, que generalmente conduce a la inhibición más alta posible. Además, la selección natural puede actuar más rápido para resistir (o desintoxicar) abundantes fitoquímicos del suelo que contra compuestos menos abundantes en el suelo.
Crecimiento - defensa
A menudo puede haber una compensación entre el crecimiento y la defensa de la planta, como la asignación de recursos para la producción de aleloquímicos (Hartley y Jones, 1997).
No hay compensación
En otros casos, tal compensación puede no existir, y un aumento en la producción de agentes alelopáticos también mejora la capacidad competitiva de la planta (Siemens et al. 2002).
Diferencia beneficios - costos de la producción
En ambos casos, las especies donantes pueden ser incapaces de optimizar su efecto alelopático maximizando la exudación. En cambio, para lograr un ESS, las especies donantes deben maximizar la diferencia entre los beneficios y los costos de producir el efecto alelopático.
Diferencia beneficios - costos de la resistencia
Obviamente, las plantas receptoras tienen el objetivo opuesto: maximizar la diferencia entre los beneficios y los costos de resistir el efecto alelopático.De hecho, para ser un ESS, los beneficios de la resistencia deberían ser más altos que los costos en cualquier especie objetivo.
Conclusiones
Es fácil imaginar cómo una planta puede beneficiarse de la capacidad de liberar compuestos que dañan otras plantas a propósito. Una planta podría obstaculizar e incluso matar otras plantas para que éstas nunca tuvieran un cambio que comenzara a limitar su crecimiento (véase Wardle et al. 1998). Una planta podría evitar el crecimiento de otras plantas para que no pudieran consumir recursos limitantes de manera tan eficiente (ver Weih y Karlsson, 1999). Una planta podría advertir a otras plantas y, por lo tanto, "aconsejarlas" que crezcan en otro lugar (ver Mahall y Callaway, 1992). Dado que la producción de metabolitos secundarios para un propósito puede reducir la asignación de recursos en otros lugares, una planta se beneficiaría aún más si fuera capaz de concentrar la liberación de aleloquímicos temporal y espacialmente. La inhibición podría tener lugar en un momento especial cuando las plantas objetivo son más sensibles, o podría concentrarse en parches de nutrientes para que las plantas competidoras no pudieran utilizar los nutrientes de la manera más eficiente. Actualmente, sin embargo, gran parte de la investigación se ha concentrado en considerar si la alelopatía existe como un fenómeno natural. Dado que esto ya no es de la mayor importancia, la próxima investigación ecológicamente orientada sobre la alelopatía se concentrará en determinar la importancia ecológica y evolutiva de las interacciones alelopáticas en comparación con otros procesos evolutivos que afectan a las plantas. Por lo tanto, todavía vale la pena estar de acuerdo con la opinión de Muller (1969): “Los agrónomos y horticultores han estado más dispuestos que los ecologistas a considerar la alelopatía como un fenómeno significativo. Esta disparidad ahora está disminuyendo a medida que los ecologistas encuentran evidencia cada vez más efectiva de alelopatía. Una influencia importante en este desarrollo ha sido la reciente disponibilidad de poderosas técnicas analíticas químicas ... Por lo tanto, la alelopatía ha surgido como un fenómeno natural que debe considerarse en cualquier intento de comprender ese intrincado patrón llamado "proceso ecológico".
¿Tú qué concluyes?